膵液に含まれる二大消化酵素であるトリプシンとキモトリプシンは、どのような違いがあるのでしょうか?基質特異性、活性化機構、最適pH、臨床応用まで詳しく解説します。知っておくべき重要な相違点とは何でしょうか?
セリンプロテアーゼ キモトリプシンの基質特異性と生理機能
セリンプロテアーゼ キモトリプシンの構造的特徴、触媒メカニズム、生体内での役割について医療従事者向けに詳しく解説。芳香族アミノ酸への特異性の仕組みとは?
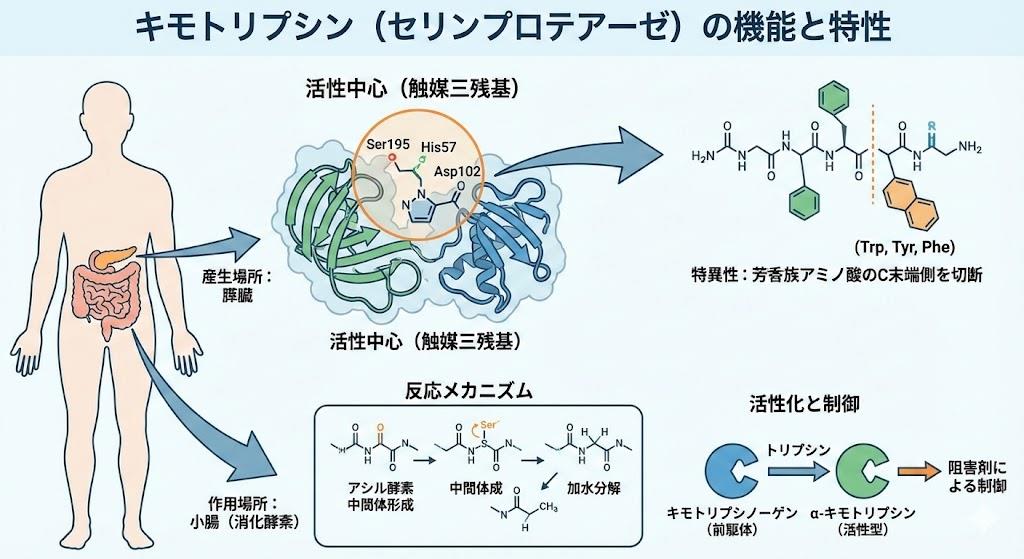

キモトリプシンは、分子量約25kDaのセリンプロテアーゼファミリーに属するエンドペプチダーゼです。この酵素の最も特徴的な構造は、キモトリプシンフォールドと呼ばれるタンデムβ-バレル構造を持つことです。
参考)https://pmc.ncbi.nlm.nih.gov/articles/PMC7615277/
活性中心には触媒三残基(catalytic triad)と呼ばれる3つのアミノ酸残基が配置されています:
参考)301 Moved Permanently
この触媒三残基の配置は、セリンプロテアーゼファミリー全体で高度に保存されており、共通の触媒メカニズムの基盤となっています。キモトリプシンの活性中心周辺には8つの表面ループが存在し、これらのループの長さや配置の違いが各セリンプロテアーゼの基質特異性を決定しています。
参考)301 Moved Permanently
特に重要なのは、S1ポケットと呼ばれる基質結合部位の構造です。キモトリプシンのS1ポケットは疎水性の深い空洞を形成しており、この構造が芳香族アミノ酸の側鎖を選択的に認識する基盤となっています。
参考)https://www10.showa-u.ac.jp/~biolchem/H20-P2Enzyme.pdf
キモトリプシンの最も顕著な特徴は、芳香族アミノ酸に対する高い基質特異性です。具体的には、フェニルアラニン(Phe)、チロシン(Tyr)、トリプトファン(Trp)のC端側ペプチド結合を選択的に加水分解します。
参考)キモトリプシン - Wikipedia
この基質特異性は、以下の分子機構によって実現されています。
疎水性ポケットによる認識
キモトリプシンのS1ポケットは深い疎水性空洞を形成しており、芳香族側鎖がこの空洞に結合することで基質-酵素複合体が安定化されます。この相互作用により、芳香族アミノ酸を含むペプチドが優先的に結合し、切断されるのです。
他のセリンプロテアーゼとの比較
同じセリンプロテアーゼファミリーでも、S1ポケットの構造の違いにより基質特異性が大きく異なります:
この比較から、キモトリプシンの疎水性ポケットがいかに芳香族アミノ酸認識に特化しているかが理解できます。
キモトリプシンによる加水分解反応は、精巧な二段階機構により進行します。この機構は、セリンプロテアーゼファミリー共通の特徴でもあります。
第一段階:アシル化反応
第二段階:脱アシル化反応
この反応機構において、Asp-102は電荷リレー系の一部として機能し、His-57の適切なプロトン化状態を維持する役割を果たしています。
反応の全体的な速度は、基質の結合親和性と触媒効率の両方によって決定されます。キモトリプシンの場合、芳香族アミノ酸を含む基質に対して特に高い触媒効率を示します。
キモトリプシンは、他の消化酵素と同様に、不活性な前駆体として合成・分泌される安全機構を持っています。この過程は「チモーゲン活性化」と呼ばれ、膵臓の自己消化を防ぐ重要なシステムです。
キモトリプシノーゲンの構造
膵臓の腺房細胞で合成されるキモトリプシノーゲンは、245個のアミノ酸からなる不活性前駆体です。この前駆体では、活性中心の形成が不完全で、プロテアーゼ活性を示しません。
段階的活性化過程
この段階的活性化により、膵臓内では不活性状態を保ちながら、必要な部位で効率的に活性化されるシステムが構築されています。
遺伝的背景
ヒトキモトリプシンの遺伝子は第16染色体のq23-q24.1領域に位置するCTRB遺伝子によってコードされています。この遺伝子の変異は、慢性膵炎や膵癌のリスク因子として研究されています。
キモトリプシンは消化系における中核的な役割を担っているだけでなく、様々な生理学的プロセスや病態に関与しています。
消化における役割
膵液中のキモトリプシンは、最適pH 8-9の弱塩基性条件下で最高活性を示します。十二指腸内でのタンパク質消化において:
皮膚における機能
興味深いことに、キモトリプシン様プロテアーゼは消化管以外でも重要な機能を果たしています。表皮のケラチノサイト(角化細胞)でも産生され:
参考)https://www.sccj-ifscc.com/library/glossary_detail/411
阻害剤との相互作用
セルピン(セリンプロテアーゼ阻害剤)は、キモトリプシンの活性を厳密に制御する重要な調節因子です。セルピンは独特な阻害機構を持ち:
参考)セルピン - Wikipedia
病理学的関連性
キモトリプシンの異常は多様な疾患と関連しています。
現在、キモトリプシンを標的とした治療薬の開発も進められており、特にプロテアーゼ阻害剤は抗炎症薬や皮膚治療薬として有望視されています。
参考)https://patents.google.com/patent/JP2011511766A/ja
ブラックキャップ [12個入] ゴキブリ駆除剤 固形物 食いつき2.5倍! 置いたその日から効く 防除用医薬部外品 【Amazon.co.jp限定】