骨芽細胞の構造や機能、最新研究から分化誘導を活用した再生医療までを詳細に解説。骨代謝の鍵を握る細胞の可能性を探るとともに、臨床応用の展望とは?
TRAP 破骨細胞特異的遺伝子と骨吸収の分子機構
破骨細胞に特異的に発現するTRAP遺伝子の構造と機能、さらに骨代謝における重要性について最新の研究知見を解説します。臨床応用への可能性は広がっているのでしょうか?

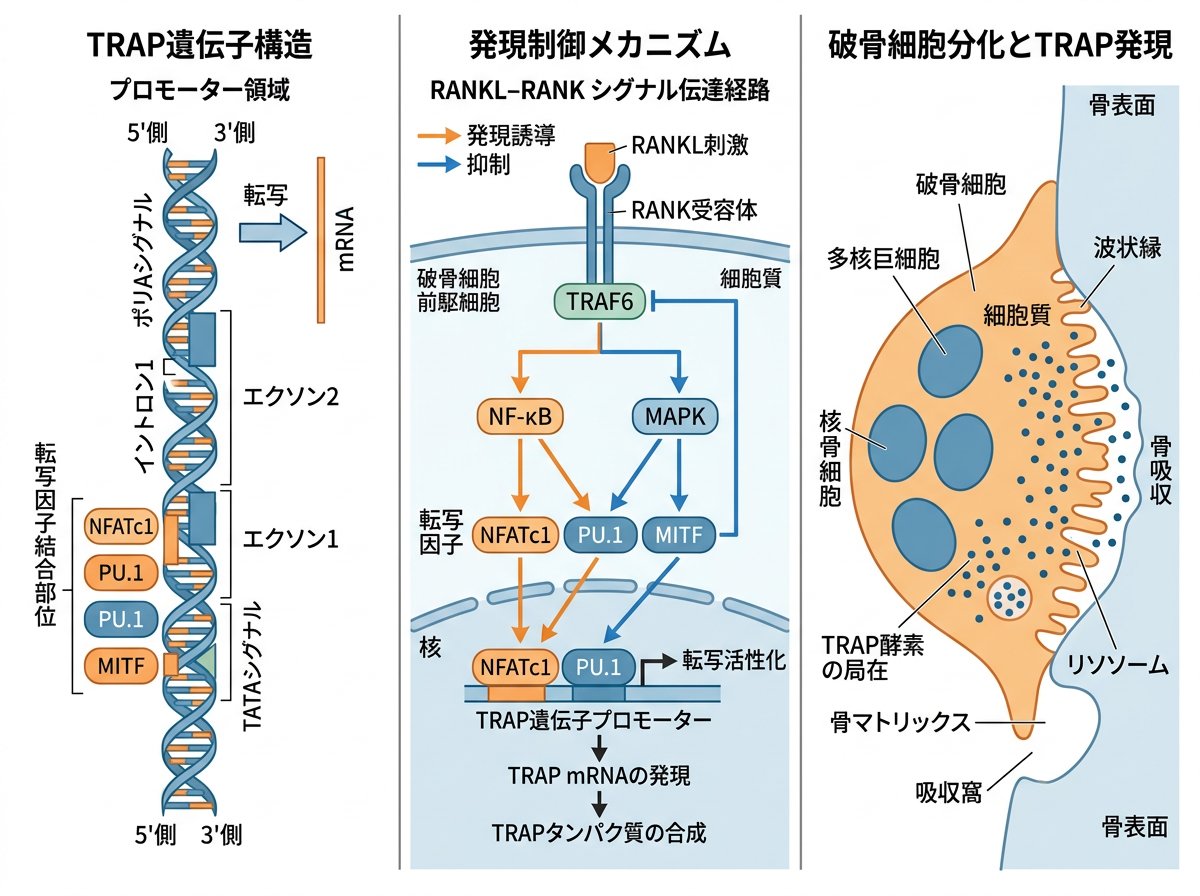
TRAP(Tartrate-Resistant Acid Phosphatase)は、破骨細胞に高度に特異的に発現する遺伝子として知られており、その遺伝子産物は酒石酸耐性酸ホスファターゼとして機能します。TRAP遺伝子は、ACP5(Acid Phosphatase 5)とも呼ばれ、ヒトでは19番染色体上に位置しています。
TRAP遺伝子のプロモーター領域には、いくつかの重要な転写因子結合部位が同定されています。特に注目すべきは、PU.1、MITF(Microphthalmia-associated Transcription Factor)、NFATc1(Nuclear Factor of Activated T-cells c1)などの転写因子結合領域です。これらの転写因子はいずれも破骨細胞分化のカスケードにおいて重要な役割を果たしています。
TRAP遺伝子の発現調節には、複数のシグナル伝達経路が関与しています。
興味深いことに、TRAP遺伝子のプロモーター領域にはPAX6の結合サイトも存在し、PAX6はNFATc1によるTRAP遺伝子の転写活性化を抑制することが報告されています。これは破骨細胞分化の負の制御機構として機能していると考えられます。
破骨細胞分化過程におけるTRAP遺伝子の発現パターンは、時間的に厳密に制御されています。造血幹細胞から破骨細胞前駆細胞への分化初期には、M-CSF(Macrophage Colony-Stimulating Factor)が重要な役割を果たしますが、この段階ではTRAP遺伝子の発現は低レベルにとどまっています。
RANKL刺激による破骨細胞分化誘導過程では、以下のような時間的制御が観察されます。
特に注目すべきは、NFATc1がTRAP遺伝子の発現を直接的に制御している点です。研究によれば、NFATc1はc-Fosの下流で機能し、c-Fosノックアウトマウスに見られる大理石骨病(破骨細胞欠損による骨吸収障害)の表現型はNFATc1の過剰発現によって回復することが示されています。
また、Jdp2(Jun Dimerization Protein 2)というAP-1ファミリーに属する転写因子もTRAP遺伝子の発現制御に関与しています。Jdp2ノックアウトマウスではRANKLによるc-Fosの活性化は正常に生じるものの、TRAPやカテプシンKの誘導が起こらないことから、Jdp2はc-Fosの下流で機能し、破骨細胞特異的遺伝子の発現誘導に必須であると考えられています。
TRAP遺伝子の転写制御において、NFATc1とMITFは中心的な役割を果たしています。これらの転写因子は協調的に働き、破骨細胞特異的な遺伝子発現プログラムを実行します。
NFATc1は破骨細胞分化のマスター転写因子と位置づけられています。NFATc1の活性化には二つの主要な経路が関与しています。
これらの経路が統合されることで、NFATc1は核内へと移行し、TRAP遺伝子を含む破骨細胞特異的遺伝子群の転写を活性化します。
一方、MITFは基本的にはメラニン産生細胞に発現する転写因子として知られていますが、破骨細胞においても重要な役割を果たしています。MITFには複数のアイソフォームが存在し、特にMitf-Eは破骨細胞分化に正の相関を示すことが報告されています。
研究によれば、PU.1とMITFは相乗的にTRAP遺伝子の転写を活性化することが知られています。また、NFATc1やc-Fosなどの転写因子もこの過程に関与しています。
TRAP遺伝子の転写活性化における詳細なメカニズムとしては、以下のようなモデルが提案されています。
興味深いことに、TGF-β(Transforming Growth Factor-β)はマウスとイヌでは異なる効果を示し、マウスの破骨細胞分化を促進する一方、イヌでは抑制することが報告されています。これらの種差はMitf-Eの発現パターンの違いと関連している可能性があります。
破骨細胞分化におけるTGF-βの影響とMitf-Eの役割に関する研究
TRAPは単なるマーカー遺伝子ではなく、破骨細胞の骨吸収機能において重要な役割を果たしています。TRAP遺伝子がコードする酒石酸耐性酸ホスファターゼは、破骨細胞の吸収窩において骨基質タンパク質の分解に関与しています。
破骨細胞による骨吸収の過程は以下のように進行します。
TRAP酵素は特に、骨基質のリン酸化タンパク質の脱リン酸化に関与しており、これによってカテプシンKなどのプロテアーゼによる分解を促進すると考えられています。また、TRAPは活性酸素種(ROS)の生成にも関与し、これが骨基質タンパク質の分解を促進する可能性も示唆されています。
TRAP活性の重要性は、TRAP遺伝子をノックアウトしたマウスの表現型からも支持されています。TRAP欠損マウスでは、骨吸収の障害による軽度の大理石骨病様の表現型が観察されています。
臨床的には、血清TRAP 5b(破骨細胞由来のTRAPアイソフォーム)は骨吸収マーカーとして使用されており、骨粗鬆症や悪性腫瘍の骨転移などの診断や治療効果のモニタリングに役立っています。
近年、光遺伝学(オプトジェネティクス)技術を用いた破骨細胞機能の時空間的制御に関する研究が進展しています。これは従来の薬物や遺伝子工学的手法とは異なり、光照射によって特定の遺伝子発現や細胞機能を非侵襲的かつ高精度に制御する革新的なアプローチです。
東京医科歯科大学の研究グループは、光を用いて破骨細胞の分化を誘導し、成熟した破骨細胞を生み出す光遺伝学ツールの開発に成功しました。この技術を用いることで、光照射をコントロールすることにより骨吸収を空間的に制御することが可能になりました。
この技術の破骨細胞特異的遺伝子の研究への応用として、以下のような可能性が考えられます。
この技術の臨床応用としては、骨粗鬆症や関節リウマチなどの疾患において、病的な骨吸収が起こっている特定の部位での破骨細胞の活性を選択的に抑制する治療法の開発が期待されています。
また、TRAP遺伝子のプロモーターを利用して、破骨細胞特異的に光応答性イオンチャネルや光活性化型薬物放出システムを発現させる研究も進められています。これにより、破骨細胞の機能を光で直接制御することが可能になると期待されています。
破骨細胞の形態形成や融合過程についても、光遺伝学技術を用いた研究が進んでいます。TRAP遺伝子と共発現するDC-STAMP(Dendritic Cell-Specific Transmembrane Protein)やATP6V0d2などの破骨細胞融合関連遺伝子の発現を光で制御することで、破骨細胞の多核化プロセスの解明が進むことが期待されています。
東京医科歯科大学による光遺伝学を用いた骨吸収制御技術の開発についてのプレスリリース
これらの最新研究は、TRAP遺伝子を含む破骨細胞特異的遺伝子の基礎研究のみならず、骨代謝疾患の新たな治療戦略の開発にも大きく貢献するものと期待されています。
ブラックキャップ [12個入] ゴキブリ駆除剤 固形物 食いつき2.5倍! 置いたその日から効く 防除用医薬部外品 【Amazon.co.jp限定】