骨芽細胞の構造や機能、最新研究から分化誘導を活用した再生医療までを詳細に解説。骨代謝の鍵を握る細胞の可能性を探るとともに、臨床応用の展望とは?
カテプシンKと破骨細胞の酵素活性と病態
骨代謝において重要な役割を果たすカテプシンKと破骨細胞の関係性について詳しく解説します。酵素活性機序から病態への影響、最新の治療戦略まで、医療専門家が知っておくべき情報を網羅していますが、カテプシンKの新たな機能とは何でしょうか?

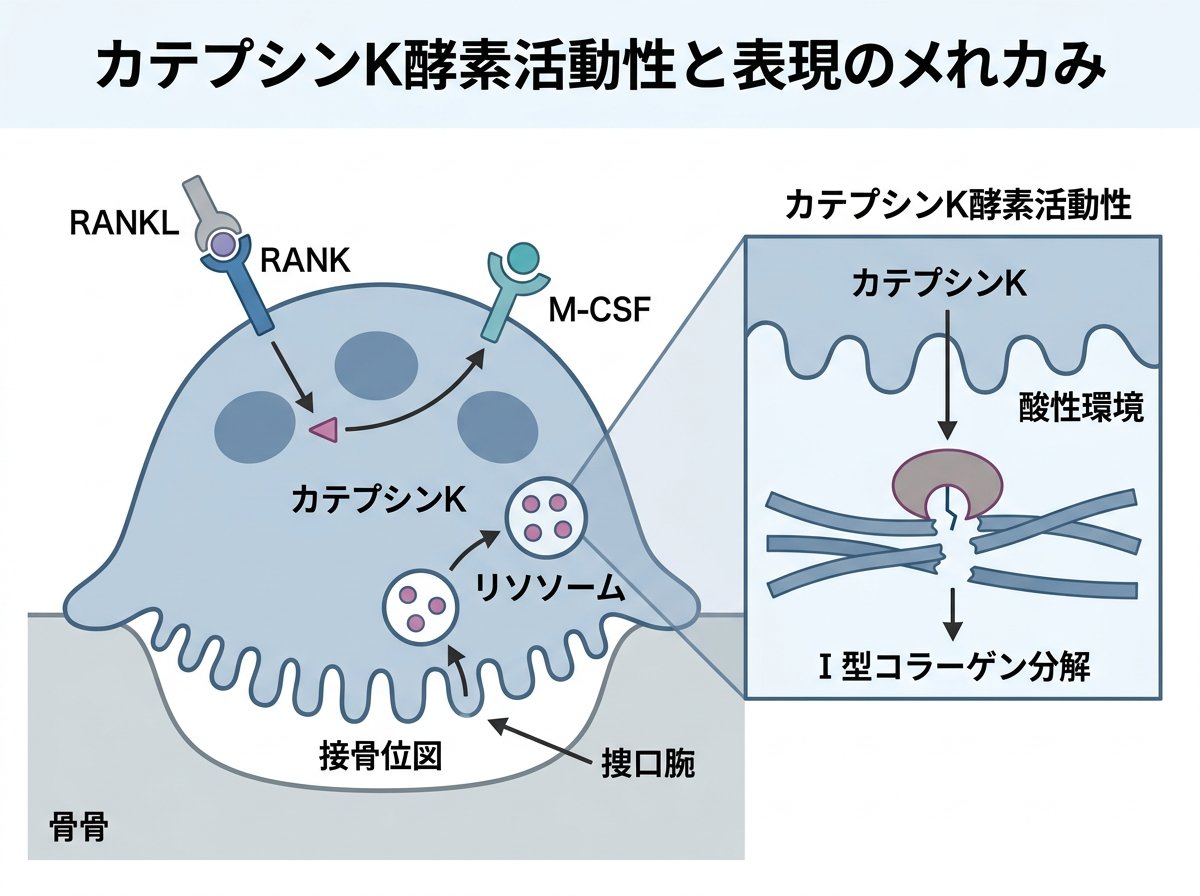
カテプシンKは、リソソームに局在するタンパク質分解酵素(システインプロテアーゼ)の一種です。ヒトでは11種類(B, C, F, H, K, L, O, S, V, W, X)のカテプシンが知られていますが、カテプシンKはその中でも特に破骨細胞に高発現している酵素として注目されています。他のカテプシンがリソソーム内で機能するのに対し、カテプシンKは破骨細胞の酸性環境である骨吸収窩に分泌され、骨基質の分解という特殊な役割を担っています。
破骨細胞は骨髄由来の単球/マクロファージ系の前駆細胞が融合して形成される多核巨大細胞です。これらの細胞は、RANKL(receptor activator of nuclear factor kappa-B ligand)などのシグナルを受けて分化します。分化した破骨細胞は、骨表面に密着して「封鎖領域」を形成し、その内部にプロトンポンプによって酸性環境を作り出します。この酸性環境下でカテプシンKは最適な活性を示し、骨の有機成分の約90%を占めるI型コラーゲンを効率的に分解します。
カテプシンKの発現は、破骨細胞の分化過程で特異的に誘導されます。この発現には転写因子NFATc1(nuclear factor of activated T cells c1)やPU.1が重要な役割を果たしています。また、カテプシンKは骨吸収の過程で、細胞内のリソソームから分泌顆粒を介して骨吸収窩へと輸送されます。この輸送過程には小胞輸送タンパク質Rab7やSynaptotagmin VIIが関与していることが明らかになっています。
破骨細胞の機能を特徴づける重要な酵素として、カテプシンKに加えて酒石酸抵抗性酸ホスファターゼ(TRAP)が挙げられます。TRAPは破骨細胞の分化マーカーとして広く用いられており、骨代謝研究において欠かせない指標となっています。
カテプシンKとTRAPは、破骨細胞の骨吸収機能において相互に補完的な役割を果たしています。カテプシンKが骨基質のコラーゲン線維を分解する一方、TRAPはリン酸化タンパク質の脱リン酸化を担当しています。特に、TRAPはオステオポンチンなどの骨基質タンパク質の脱リン酸化を通じて、破骨細胞の移動や接着を調節していると考えられています。
興味深いことに、カテプシンKとTRAPの発現パターンには相関関係が見られます。両酵素とも破骨細胞の分化過程で誘導され、活性化された破骨細胞で高レベルに発現します。また、骨粗鬆症などの骨代謝疾患では、両酵素の発現レベルが変化することが報告されています。
最近の研究では、カテプシンKがTRAPの活性化に関与している可能性が示唆されています。カテプシンKは、不活性な前駆体として合成されるTRAPを限定分解することで活性型へと変換する機能を持つと考えられています。このように、両酵素は単に並行して機能するだけでなく、互いの活性を調節する複雑なネットワークを形成していることが明らかになりつつあります。
TRAP活性化におけるカテプシンKの役割についての詳細はこちらの論文で解説されています
カテプシンK遺伝子(CTSK)の変異や欠損は、ヒトにおいて濃化異骨症(ピクノディソストーシス)という稀な常染色体劣性遺伝疾患を引き起こします。この疾患は骨硬化(骨密度の増加)、易骨折性、低身長、頭蓋顔面異常などを特徴とします。フランスの画家トゥールーズ=ロートレックがこの疾患に罹患していたことでも知られています。
ピクノディソストーシスでは、カテプシンK活性の欠如により破骨細胞による骨基質の分解が障害されます。しかし興味深いことに、破骨細胞の形成自体は正常に進行し、骨の無機成分(ハイドロキシアパタイト)の溶解も起こります。つまり、破骨細胞の分化と骨吸収窩の酸性化は正常に行われるものの、有機基質(主にコラーゲン)の分解が特異的に障害されるのです。
実験動物モデルでも同様の現象が確認されています。カテプシンK遺伝子欠損マウスでは、骨量の増加(骨硬化)が観察されますが、破骨細胞の数自体は正常かむしろ増加していることが報告されています。これは、カテプシンK欠損下では骨吸収能が低下するものの、それを補償するためにより多くの破骨細胞が形成されることを示唆しています。
また、カテプシンK欠損は骨代謝のバランスにも影響を及ぼします。通常、破骨細胞による骨吸収と骨芽細胞による骨形成はカップリングして進行しますが、カテプシンK欠損状態ではこのカップリングが部分的に解除(アンカップリング)され、骨形成が相対的に優位になることが示されています。この現象は、カテプシンK阻害薬の開発において重要な理論的基盤となっています。
さらに、カテプシンKの機能異常は加齢に伴う骨粗鬆症の病態にも関与している可能性があります。加齢とともにカテプシンKの活性調節機構に変化が生じ、過剰な骨吸収につながるという仮説も提唱されています。
骨粗鬆症をはじめとする骨代謝疾患の治療において、破骨細胞とカテプシンKは重要な治療標的となっています。特にカテプシンK阻害薬は、既存の骨吸収抑制薬とは異なるメカニズムで作用する新しいクラスの薬剤として注目されています。
従来の骨吸収抑制薬であるビスホスホネート製剤は、破骨細胞のアポトーシス(細胞死)を誘導することで骨吸収を抑制します。一方、カテプシンK阻害薬は破骨細胞の生存や分化には影響せず、骨基質分解能のみを選択的に阻害します。このような作用機序の違いから、カテプシンK阻害薬は骨吸収と骨形成のカップリングを完全には遮断せず、骨形成を維持したまま骨吸収を抑制できる可能性があります。
オダナカチブ(MK-0822)は最も開発が進んだカテプシンK阻害薬の一つでした。臨床試験では骨密度の増加と骨折リスクの低減が確認されましたが、脳卒中リスクの増加という安全性の懸念から開発が中止されました。しかし、この副作用はカテプシンK阻害作用とは直接関連しないと考えられており、より選択性の高い次世代のカテプシンK阻害薬の開発が進められています。
また、破骨細胞の分化や活性化を制御する別のアプローチも研究されています。RANKL抗体であるデノスマブは、破骨細胞前駆細胞の分化を阻害することで骨吸収を抑制し、骨粗鬆症治療薬として実用化されています。さらに、破骨細胞特異的な細胞表面分子やシグナル伝達経路を標的とした新規治療法の開発も進んでいます。
最近では、破骨細胞とカテプシンKに対する二重標的アプローチも検討されています。例えば、破骨細胞の分化を部分的に抑制しつつ、カテプシンK活性も阻害することで、より効果的に骨吸収を制御する戦略が提案されています。このような複合的なアプローチは、単一標的治療の限界を克服し、より効果的で副作用の少ない治療法につながる可能性があります。
骨粗鬆症の薬物療法ガイドラインにおけるカテプシンK阻害薬の位置づけについてはこちらで詳しく解説されています
カテプシンKと破骨細胞に関する研究は近年急速に進展していますが、今後さらに発展が期待される分野もあります。特に注目されているのが、カテプシンKの骨代謝以外での機能解明です。
最近の研究では、カテプシンKが脂肪組織や免疫系においても重要な役割を果たしていることが示唆されています。例えば、カテプシンKは脂肪細胞の分化や機能調節に関与しており、肥満や脂質代謝障害との関連が指摘されています。また、自己免疫疾患である関節リウマチの病態形成にも関与している可能性があり、炎症性サイトカインの制御や自己抗原の産生に影響を与えると考えられています。
破骨細胞研究においては、単一細胞解析技術の進歩により、破骨細胞の不均一性(ヘテロジェニティ)が明らかになりつつあります。破骨細胞は単一の細胞集団ではなく、起源や機能が異なる複数のサブタイプに分類できることが示唆されています。このような破骨細胞の多様性の理解は、より精密な骨代謝制御メカニズムの解明につながるでしょう。
また、骨代謝と全身の他の生理機能との連関(オステオネットワーク)の研究も進んでいます。破骨細胞が産生する因子(オステオカイン)が、エネルギー代謝や神経機能、免疫応答など全身の様々な生理機能に影響を与えることが明らかになってきました。この中でカテプシンKが果たす役割についても、今後の研究の進展が期待されます。
さらに、ゲノム編集技術や組織特異的遺伝子操作技術の発展により、より精密なモデル動物の作出が可能になっています。例えば、破骨細胞特異的にカテプシンK発現を時期特異的に制御できるモデルなどを用いた研究により、カテプシンKの機能をより詳細に解明できるでしょう。
デジタル技術の医療応用も進んでおり、AI(人工知能)を活用した骨代謝マーカーの解析や、カテプシンK阻害薬の個別化医療への応用も検討されています。例えば、骨代謝の状態やカテプシンK活性を非侵襲的に評価するバイオマーカーの開発により、より効果的な治療介入が可能になるかもしれません。
これらの研究の進展は、骨粗鬆症や関節リウマチなどの既存の疾患治療に新たな視点をもたらすだけでなく、加齢に伴う骨格系の変化や全身の代謝調節機構の理解にも貢献することが期待されます。カテプシンKと破骨細胞の研究は、単なる骨代謝研究の枠を超えて、全身の健康と疾患の理解に重要な知見をもたらす可能性を秘めているのです。
破骨細胞とカテプシンK研究の最新動向については日本骨代謝学会誌で詳しく解説されています
[指定医薬部外品] 大正製薬 新ビオフェルミンS錠 550錠 61日分整腸剤【Amazon.co.jp限定】 [乳酸菌/ビフィズス菌/フェーカリス菌/アシドフィルス菌 配合] 腸内フローラ改善 便秘や軟便に